Cristóbal Lárez Velásquez1, Maura Rojas Pirela2, Alicia Chirinos3, Luz Rojas Avelizapa4
1) Grupo de Polímeros, Departamento de Química, Facultad de Ciencias, Universidad de Los Andes. Mérida 5101 – Venezuela. Correo electrónico: clarez@ula.ve
2) Departamento de Biología, Facultad de Ciencias, Universidad de Los Andes, Mérida 5101, Venezuela: mlrojas.pirela@gmail.com
3) Departamento de Química, Universidad Politécnica Territorial Alonso Gamero, Coro–Venezuela. Correo electrónico: alibelchi@gmail.com
4) Facultad de Ciencias Biológicas y Agropecuarias. Universidad Veracruzana. km 1 Camino Peñuela–Amatlán, S/N. Peñuela, Municipio de Amatlán de Los Reyes. C.P., 94945 Veracruz, México. Correo electrónico: luzirenemx@hotmail.com
Recibido: Febrero 2019; Aceptado:Abril 2019
Texto completo (pdf)
Cita (APA)
Lárez Velásquez, C., Chirinos, A., Rojas Avelizapa, L. (2019). Nuevos retos en agricultura para los biopolímeros de quitina y quitosano. 1. Efectos beneficiosos para los cultivos. Revista Iberoamericana de Polímeros, 20(3), 118-136.
RESUMEN
En este trabajo se revisan los principales efectos observados durante la aplicación de quitina y quitosano como agentes coadyuvantes en el aumento de la producción agrícola, haciendo énfasis en los aspectos relacionados con sus efectos beneficiosos sobre el crecimiento de las plantas y en sus potenciales aplicaciones para el control de diversas plagas y enfermedades que generan grandes pérdidas en los cultivos. Se hace un esfuerzo importante para sistematizar la información sobre los diversos mecanismos de acción propuestos para que estos materiales logren la efectividad que los hacen ser tan útiles en ese campo. En ambos casos, los modos de acción considerados para organizar la amplia información existente en la literatura revisada han sido: (a) la generación del llamado efecto fertilizante, (b) la inhibición indirecta de fitopatógenos por los subproductos de su descomposición, incluyendo la generación de un factor fungistático volátil, (c) la promoción/soporte de microorganismos beneficiosos tales como antagonistas de fitopatógenos, productores de sustancias biocidas y colonizadores micorrizales, (d) sus actividades elicitoras para activar mecanismos de defensa, incluyendo la inducción de proteínas relacionadas a la patogénesis, enzimas de defensa, metabolitos secundarios y la activación de receptores específicos.
ABSTRACT
In this paper we review the main effects observed during the application of chitin and chitosan as coadjuvant agents in the increase of agricultural production, emphasizing the aspects related to their beneficial effects on the growth of plants and their potential applications for the control of various pests and diseases that generate large losses in crops. An important effort is made to systematize the information on the various mechanisms of action proposed so that these materials achieve the effectiveness that makes them so useful in this field. In both cases, the modes of action considered to organize the extensive information existing in the reviewed literature have been: (a) the generation of the so–called fertilizing effect, (b) the indirect inhibition of phytopathogens by byproducts of their decomposition, including the generation of a volatile fungistatic factor, (c) the promotion/support of beneficial microorganisms such as phytopathogen antagonists, producers of biocidal substances and mycorrhizal colonizers, (d) their eliciting activities for activating defense mechanisms, including the induction of proteins related to pathogenesis, defense enzymes, secondary metabolites and the activation of specific receptors.
INTRODUCCIÓN
El hambre es uno de los males más preocupantes en la actualidad para muchas poblaciones de seres humanos en nuestro planeta. Seguramente la situación debe ser igual de dramática en numerosas poblaciones de animales silvestres, la cual ha sido obviamente menos cubierta por los medios de información aún cuando existen campañas para evitar sus consecuencias: la extinción de muchas especies. Aunque las razones para la aparición de este mal son diversas, hay algunas similitudes en ambas. Por ejemplo, en los dos casos existe una disminución crítica en la capacidad de generación de la cantidad mínima necesaria de alimentos para sustentar una población determinada. Sin embargo, en el caso de las poblaciones humanas hay una diferencia agravante debido a que éstas, paradójicamente, aumentan su número de manera irracional, lo que ha conducido a declaraciones preocupantes sobre la existencia de una emergencia planetaria sin precedentes [1], que pareciera poder trocarse rápidamente en una emergencia planetaria instantánea [2] debido a una de sus derivaciones más perjudiciales: el calentamiento global.
Por ello, en el caso de las poblaciones humanas, el estudio de las situaciones causales permite proponer que el problema actual del hambre tiene básicamente dos fuentes primordiales: (a) el aumento exponencial de la población, y (b) la disminución a niveles críticos de la capacidad de generación de alimentos. A su vez, entre las contribuciones importantes a esta última se pudieran mencionar: (i) la disminución de las tierras aptas para cultivos, (ii) la disminución de la cantidad y calidad del agua para usos agrícolas, y (iii) las grandes pérdidas asociadas a plagas y enfermedades en los cultivos.
En la búsqueda de remediar esta situación, en alguna extensión, el hombre ha venido actuando sobre las diferentes fuentes del problema, tratando de aplicar correctivos tanto en el control del crecimiento poblacional como en el aumento de la producción de alimentos. En el primer caso el algunos países se han tomado medidas severas, tanto legales como económicas, para limitar el número de descendientes que puede tener un matrimonio, así como el desarrollo de campañas agresivas para el uso de métodos anticonceptivos, emancipación de la mujer, erradicación del machismo, etc.
En el caso de las acciones para favorecer el aumento de la producción de alimentos los esfuerzos han sido dirigidos hacia la formulación de medidas legales para evitar la desforestación y la desertificación, el desarrollo de campañas para fomentar el uso racional del agua, el uso de métodos ecológicamente limpios para el tratamiento de las aguas servidas, el empleo de fertilizantes, agroquímicos y biocidas más amigables con el ambiente. En este último punto, la investigación actual se ha venido desplazando hacia el uso de materiales biodegradables y cuyas fuentes sean de preferencia renovables. En tal sentido, los biopolímeros conocidos como quitina (poli–N–acetilglucosamina), y su derivado quitosano (obtenido por desacetilación de la quitina en porcentajes mayores al 50%, ver figura 1), han logrado un lugar preponderante en el desarrollo de aplicaciones relacionadas con el tratamiento de aguas [3], control de la humedad de suelos [4], liberación inteligente de fertilizantes [5], estimulante del crecimiento [6], inductor de mecanismos de defensa en plantas [7], antimicrobiano [8], etc. Su utilidad en esta lucha es tal que deberían incluirse también sus aplicaciones relacionadas con el control del aumento de la población debido a sus potenciales aplicaciones para la fabricación de preservativos [9,10] y para la liberación controlada de anticonceptivos [11] y espermaticidas [12], incluyendo las expectativas de crear un anticonceptivo femenino no hormonal sin efectos colaterales [13]. Más aún, si la castración quirúrgica de progenitores irresponsables llegase a ser una opción, seguramente el quitosano podría hacer una contribución importante pues desde hace mucho tiempo han sido reconocidas sus bondades en la aceleración de la cicatrización de heridas [14,15].
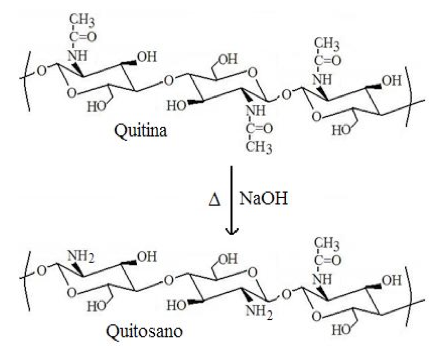
En este trabajo¸ que es la primera parte de una serie de tres artículos, se detallarán los principales efectos observados durante la aplicación de quitina y quitosano en sistemas agrícolas. Se abordan las principales aplicaciones de la quitina y el quitosano como agentes para coadyuvar en el aumento de la producción de alimentos, haciendo énfasis en los aspectos relacionados con sus efectos beneficiosos sobre el crecimiento de las plantas y en sus aplicaciones para el control de diversas plagas y enfermedades que generan grandes pérdidas en los cultivos. Se hace un esfuerzo importante para tratar de organizar la información sobre los diversos mecanismos de acción propuestos para que estos materiales logren la efectividad que los hacen ser tan útiles en ese campo. En los dos trabajos futuros se discutirán los principales mecanismos propuestos para justificar su acción directa sobre fitopatógenos (parte 2) y se detallarán los muy diversos usos que se han diseñado para estos materiales en la dosificación de fertilizantes y/u otras sustancias con actividad biológica (parte 3).
La quitina y sus efectos beneficios en plantas
Los efectos beneficiosos de la quitina en las prácticas agrícolas son conocidos desde mediados del siglo XX, cuando se comenzó a observar que algunas plantaciones cultivadas en suelos enmendados con estos materiales quitonosos mostraban un decrecimiento en ciertas enfermedades asociadas a hongos [16,17]; sin embargo, también se han reportado efectos negativos y resultados contradictorios. Así, por ejemplo, se reportó tempranamente que las enmiendas con quitina resultan en la aparición de cierta fitotoxicidad, la cual ha sido asociada a la estrecha relación N/C (6/4) que deriva de la estructura de la quitina y que resulta en un comportamiento similar al observado en otras enmiendas nitrogenadas [18]; la combinación con otros materiales, como por ejemplo desechos celulósicos, ha sido una de las vías sugeridas para mitigar el bajo contenido de carbono de las enmiendas que contienen solo quitina [19]. Otra de las causas que se ha sugerido para justificar estos efectos fitotóxicos es la alteración del pH del suelo causada por la descomposición de la quitina [20]. En ambos casos, los efectos fitotóxicos parecieran poder ser minimizados con una humectación a fondo antes del transplante [21].
Por otro lado, entre los resultados contradictorios que se han reportado están los de Mauer y Baker, quienes observaron que la quitina y la lignina aplicados separadamente no disminuyen la pudrición de la raíz del frijol pero son efectivas cuando se emplean juntas [22]; y los de Culbreath et al. mostrando que algunas enmiendas de quitina favorecieron la formación de agallas en las raíces de calabacín, por M. arenaria, en lugar de disminuirlas [23].
En la actualidad se han propuesto diversos mecanismos para explicar las rutas a través de las cuales los materiales quitinosos favorecen el desarrollo de los cultivos, incluyendo algunas que se relacionan con la inhibición del crecimiento de fitopatógenos. A continuación se resumen éstos [24]:
Ocurrencia de un efecto fertilizante. Los productos de la descomposición de la quitina en el suelo [25], tales como amonio y sus derivados, pueden constituirse en una fuente lenta pero sostenida de nitrógeno, cuya duración puede llegar incluso a los tres meses [26]. Estas circunstancias promueven el desarrollo de plantas más saludables y menos susceptibles al ataque de patógenos, como se observó en el rápido reverdecimiento de plantas de celery luego de su transplante en lotes enmendados con quitina, aún cuando ni el tamaño individual de las plantas ni el peso fresco de los tallos comercializables fueron significativamente diferentes en comparación a los obtenidos sin quitina, pero si el rendimiento total debido a la reducción en la incidencia del amarillamiento por Fusarium [27].
Inhibición indirecta de fitopatógenos por los subproductos de su descomposición. La actividad fungicida de algunos productos volátiles generados durante la descomposición de la quitina en el suelo ha sido vinculada al llamado factor fungistástico volátil, específicamente al desprendimiento de amonio [28] aunque algunos de sus alquil–derivados (metilamina, etilamina) no pueden ser completamente descartados [29]. Es importante considerar que el tipo de microorganismos presentes en el suelo es un factor clave en la liberación de diferentes volátiles, habiéndose comprobado que algunos microorganismos específicos son responsables por la liberación de una amplia variedad de compuestos volátiles [30,31].
Promoción/soporte de microorganismos beneficiosos
Promoción del crecimiento de microorganismos antagonistas. La presencia de materiales quitinosos puede cambiar la microflora del suelo para aumentar la población de antagonistas (los cuales compiten por alimento y espacio, hiperparasitismo, lisis de las hifas, etc.) a patógenos que atacan las raíces. Una estrategia para el control de este tipo de patógenos consiste en añadir enmiendas de quitina que contienen un antagonista adecuado, como por ejemplo el uso de enmiendas de quitina conteniendo Pseudomonas aeruginosa en la supresión de Macrophomina phaseolina, Rhizoctonia solani, Fusarium oxysporum y Fusarium solani en las raíces de pimientos (Capsicum annuum L.) [32]. Otros estudios han demostrado que la quitina también puede ser efectiva para disminuir el empleo de fungicidas químicos cuando es aplicada como un sustrato foliar para favorecer poblaciones bacterianas nativas con actividad lítica sobre patógenos que atacan las partes aéreas de las plantas, como en el control del hongo Mycosphaerella fijiensis Morelet, causante de la sigatoka negra en plantaciones de bananas (Musa paradisiaca L.) y plátanos (Musa acuminata Colla), mediante aplicaciones foliares que contienen quitina coloidal y que permite disminuir entre 43–46% la aplicación de fungicidas convencionales cuando se aplican rotativamente en forma de aerosol [33].
Promoción/soporte de microorganismos que producen biocidas. Los cuales operan debido a la actividad antimicrobiana de sus metabolitos secundarios, que incluyen enzimas líticas de las paredes celulares y otros tipos de compuestos con actividad biocida.
Biocidas enzimáticos: aunque no siempre la actividad quitinolítica de una enzima está relacionada directamente con su actividad fungicida [34], la estimulación del crecimiento de microorganismos que producen este tipo de enzimas es uno de los mecanismos más frecuentemente propuestos para explicar la inhibición de hongos fitopatógenos [35,36]. La actividad quitinolítica de las enzimas producidas por algunos de estos microorganismos ha sido claramente demostrada por Malathi y Wiswanathan durante estudios sobre biocontrol del hongo causante de la raíz roja de la caña (Colletotrichum falcatum), usando tanto hongos (Trichoderma spp.) como bacterias (Pseudomonas fluorescent) [37]. Usando un filtrado de sus cultivos, libres de células y a diferentes concentraciones, establecieron que el crecimiento del fitopatógeno es inhibido con una eficacia que aumenta cuando la concentración del filtrado aumenta; además, confirmaron que la quitina tiene influencia en la supresión del patógeno.
Biocidas no enzimáticos: durante el crecimiento de P. aeruginosa en un medio que contenía desechos de conchas de mariscos, San–Lang et al. aislaron un complejo polisacárido, que denominaron pafungin, quizás en alusión al nombre inicial dado por Braconnot a la quitina (fungina). Dicho complejo resultó efectivo contra las 39 cepas de hongos ensayados [38]. Similarmente, durante el crecimiento de Bacillus subtilis en un medio que contenía quitina se reportó que los compuestos no enzimáticos producidos mostraron actividad fungicida contra F. oxysporum [39].
Estímulo del crecimiento de micorrizas. Los materiales quitinosos, como por ejemplo algunos residuos procedentes de conchas de mariscos, también han demostrado un efecto estimulante sobre la colonización de fresas (Fragaria ananassa) por hongos micorrizógenos arbusculares (HMA) [40]. Por otra parte, Staehelin et al. han demostrado que los lipooligosácaridos derivados de la sustitución por ácidos grasos del grupo N–acetilo en los extremos no reductores de la quitina son moléculas bioactivas como señalizadoras [41], las cuales son secretadas por Rhizobia (conocidos como factores Nod) durante el establecimiento de la simbiosis legumbre/Rhizobia [42]. Por ello, se podrían adicionar enmiendas con oligómeros de quitina, o mejor aún, con quitosanos N–sustituidos con ácidos grasos [43], los cuales podrían actuar como precursores para estos metabolitos. De igual manera, se ha mostrado que la adición de quitina a sustratos de cultivos basados en suelos arenosos estimula la colonización de las raíces, el crecimiento de micelio extra–radical (MER) y la producción de esporas de HMA cuando las plantas hospedadoras fueron Allium ampeloprasum, Plantago lanceolata and Lactuca sativa [44]. Similarmente, también se pudo observar que la esporulación de HMA se vio estimulada cuando, en vez de quitina, se usó micelio de F. oxysporum autoclavado (el cual contiene quitina). Ahora bien, en un contexto más amplio, es importante caer en cuenta que la promoción de microorganismos que favorecen la infección micorrizal puede generar resultados similares, e indistinguibles, de aquellos atribuibles a un efecto fertilizante directo; por ello, la determinación de cual contribución es la que se está observando en un determinado sistema representa un reto interesante para abordar.
Actividad como elicitor (estimulante de mecanismos de defensa). Algunos fragmentos oligómericos de la quitina son reconocidas por las plantas como patrones moleculares asociados a microbios (PMAMs), especialmente a hongos, induciendo respuestas inmunes en ellas. Algunas de las reacciones de protección inducidas por la quitina y sus oligómeros han quedado bien establecidas. Entre éstas se pueden mencionar: la deposición de callosa en las partes maduras de las raíces de Arabidopsis [45]; el incremento en la actividad de enzimas asociadas a la lignificación en cultivos de células de zanahoria [46]; lignificación de heridas en hojas de trigo [47]; activación de enzimas relacionadas a la lignificación en hojas de soya [48], expresión de genes asociados a mecanismos de defensas [49]; producción de especies reactivas a oxígeno en células de arroz [50]; activación de proteínas kinasas activadas por mitogeno en plantas como Arabidopsis [51].
Por otro lado, también se han desarrollado diversos estudios relacionados con los receptores de quitina en las plantas, entre los cuales merecen destacarse los siguientes: el aislamiento en los rizomas de la ortiga (Urtica dioica) de una pequeña lectina que exhibe especificidad para unir quitina [52], encontrándose posteriormente que ésta también posee actividad antifúngica, con un mecanismo de acción que difiere del de las quitinasas [53]; el reporte de la percepción específica de concentraciones sub–nanomolares de quitina por células de tomate, estableciéndose que los oligómeros de 4 unidades de N–acetil–D–glucosamina tienen un efecto elicitor1 significativo a concentraciones del orden de los 10 pM[54]; Shibuya et al. propusieron por primera vez en 1993 la existencia de una proteína que une elicitores de quitina con alta afinidad [55a], obtenida a partir de suspensiones derivadas de cultivos de células de arroz, luego, en 1997 propusieron que ésta es una glicoproteína de la membrana plasmática conteniendo dos LysM extracelulares que unen quitina directamente [55b] y, finalmente, en 2006 fue aislada, caracterizada y denominada CEBiP (Chitin Elicitor–Binding protein) [55c]; también se ha descrito un receptor tipo kinasa denominado CERK1 (Chitin Elicitor Receptor Kinase), el cual es esencial en la señalización de la quitina como elicitor, como una proteína que contiene tres LysM en el dominio extracelular y una proteína básica Serina/Treonina con actividad kinasa en el dominio intracelular [56], apuntando a que este tipo de receptores juega un papel crítico en la detección de PMAMs en las plantas.
1
Efecto elicitor: En botánica, elicitor es utilizado para referirse a un grupo diversos de compuestos estructurales (extrínsecos o aportados) que actúan como moléculas señalizadoras cuando existe peligro. Dichas moléculas se unen a proteínas receptoras especiales ubicadas en las membranas celulares de las plantas generando una respuesta de defensa de la planta frente a diferentes patógenos (insectos, hongos o bacterias) o ante el daño mecánico producido por los herbívoros. Actualmente se clasifican en dos grandes grupos, patrones moleculares de daño asociado (DAMPs, por sus siglas en inglés) y patrones moleculares asociados a herbívoros (HAMP, por sus siglas en inglés).
En este contexto, algo sumamente interesante es que en esta dinámica guerra química natural los fitopatógenos también mueven sus piezas, desarrollando moléculas específicas, denominadas efectores2, que se unen a los PMAMs para “camuflarlos” a los receptores de las plantas; se ha demostrado que estos efectores son arreglos moleculares similares a los de los receptores en las plantas, como por ejemplo los LysM [57].
2
Los efectores son células nerviosas que ejecutan respuestas ante los estímulos que reciben. Todas las células de un animal tienen que responder de forma coordinada. Existen células especializadas (efectoras) en elaborar respuesta, la secreción de sustancias y el movimiento.
Efectos beneficiosos del quitosano en agricultura. El quitosano, además de poder generar efectos similares a los producidos por la quitina en sus aplicaciones agrícolas, guarda un as bajo la manga: su solubilidad en medio acuoso. Esta solubilidad le permite, entre otras cosas, tener un mayor control sobre su dosificación (solo o en mezclas), la preparación de diversas formas de aplicación (soluciones, emulsiones, hidrogeles, nanopartículas, películas, etc.) y, en general, una mejor manipulación. En esta sección, se detallarán los principales efectos observados durante la aplicación de quitosano en sistemas agrícolas, tratando en lo posible de seguir el mismo esquema presentado en la parte anterior para los mecanismos de acción de la quitina previamente reportados [24]. Una visión amplia de los modos de acción propuestos para el quitosano, y sus interrelaciones, se puede apreciar de manera gráfica en la Figura 2.
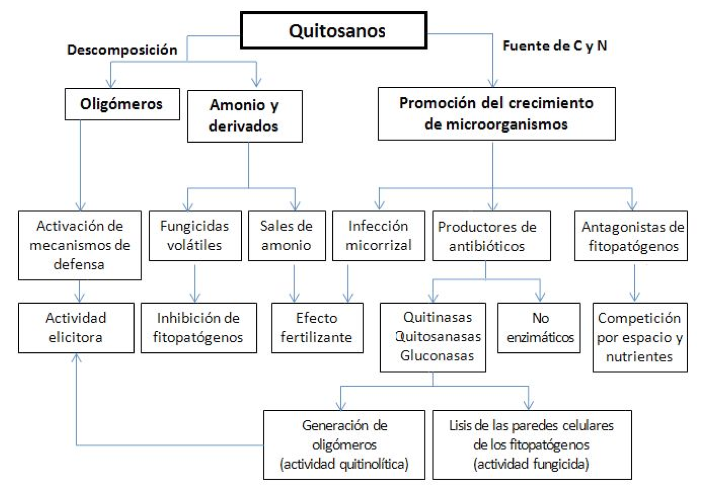
Efectos beneficiosos del quitosano sobre las plantas. Efecto fertilizante del quitosano. Se ha reportado que el uso de un suelo enmendado con quitosano en polvo (1% p/p) favoreció notoriamente tanto el reverdecimiento como el desarrollo de plantas florales transplantadas, en comparación al suelo enmendado con fertilizantes inorgánicos (con el mismo contenido de nitrógeno) y también al suelo sin enmienda [58]. Estos resultados parecen apuntar a que el quitosano estimula el desarrollo de mecanismos para una mayor captación de nutrientes nitrogenados, incluyendo el desarrollo de un mayor sistema radicular, como se ha observado en diversos cultivos tales como maíz (Zea maiz) [59] y calabacín (Cucurbita pepo), entre otros. En este último caso, el quitosano no solo favoreció la germinación in vitro más temprana [60] sino que además se pudo observar un mayor desarrollo del sistema radicular, con la aparición de un número mayor de raíces seminales (ver Figura 3) [61].

Inhibición indirecta de fitopatógenos por los subproductos de descomposición del quitosano. Aunque no se han encontrado estudios reportados sobre efectos en fitopatógenos de un hipotéticos factor fungistático volátil asociado a la degradación del quitosano en el suelo, sí hay estudios donde se han detectado subproductos volátiles como CO2 y NH3 [62], con lo cual esta posibilidad permanece abierta. Probablemente sea más difícil, en comparación con la quitina, encontrar microorganismos en el suelo que degraden quitosano debido a su escasa distribución en ambientes naturales; sin embargo, desde hace un buen tiempo se conoce que está presente en algas verdes de la especie Chlorella [63]y en las paredes celulares de algunos hongos pertenecientes a la clase Zygomicetes, como el Mucor rouxii, que lo poseen en sus paredes celulares (junto con quitina)[64]; este tipo de hongos se caracterizan por producir enzimas desacetilasas (que convierten las unidades N–acetil–glucosamina de la quitina en las unidades glucosamina del quitosano), las cuales pudieran favorecer el desarrollo de microorganismos que usan quitosano como fuente de C y N, degradándolo mediante enzimas denominadas quitosanasas.
Promoción/soporte de microorganismos beneficiosos. En algunos tipos de suelos se ha encontrado que el quitosano puede alterar la microflora promoviendo el crecimiento de actinobacterias (Streptomyces y Kitasatospores), las cuales son conocidas por producir quitosanasas[65]. A continuación se presentan algunos ejemplos específicos del tipo de efectos sobre otros microorganismos que puede ser asociado a la presencia del quitosano en cultivos.
Promoción/soporte de antagonistas: una interesante situación en este aspecto ha sido descrita para el control por Lysobacter enzymogenes, en un medio hidropónico, del patógeno Pythium aphani–dermatum, causante de la pudrición de la raíz del pepino. Los tratamientos con quitosano solo y con L. enzymogenes solo no fueron efectivos en la disminución de la enfermedad; sin embargo, el tratamiento fue exitoso cuando se aplicaron juntos; se ha propuesto que en este caso el quitosano puede actuar de dos maneras: como fuente de C y N para favorecer el desarrollo del antagonista y como inductor de la expresión de: (a) enzimas que degradan el quitosano de partida (se supone que generando oligómeros con mayor actividad biocida hacia el patógeno), (b) otras proteínas que favorecen un mayor control de la enfermedad[66].
Promoción del desarrollo de microorganismos que producen biocidas: aunque la información sobre la producción de quitosano en la naturaleza es escasa se ha demostrado que en algunos suelos costeros existen bacterias capaces de degradar quitina, para convertirla en quitosano y sus oligómeros, por vía enzimática usando N–acetil–glucosaminidasa y quitinasas[67]. Este hecho genera la posibilidad de encontrar microorganismos que usen quitosanasas para degradar el quitosano producido en tales entornos, con las consecuencias positivas que pudieran derivarse de ello para las plantas cultivadas bajo esas condiciones. Entre los efectos positivos que pudieran observarse estarían el efecto elicitor que pueden generar los oligómeros de quitosano y su actividad biocida contra fitopatógenos, además de la propia actividad biocida de las quitosanasas. Por otro lado, es importante recordar que hay plantas que pueden activar este tipo de mecanismos sin necesitar un elicitor químico, como lo demuestran los resultados del estudio con brotes de bambú, donde se ha observado un incremento en la actividad de quitosanasas cuando éstos son almacenados a temperatura ambiente, apuntando hacia la activación de mecanismos de defensa para generar estas enzimas, las cuales pueden prevenir el ataque de hongos que contengan quitosano[68]. La profundización en el estudio del rol de las quitosanasas, en conjunto con sus productos los quitooligómeros, en los mecanismos de defensa de las plantas es un campo virgen para la investigación, tanto académica como de aplicaciones, en agricultura.
Estimulante de la interacción rhizobium/planta hospedadora: se ha señalado que el inicio de la interacción rhizobium/planta para formar los nódulos fijadores de nitrógeno en las raíces de las plantas es elicitado por lipoquitooligómeros específicos de cada planta denominados factores Nod[69a]. Estos factores Nod pueden generar cambios drásticos en la formación de los nódulos cuando su estructura química es cambiada mediante sustituciones específicas[69b]. También se ha mostrado que los hongos micorrizales arbusculares usan este mismo tipo de moléculas para iniciar su relación simbiótica con plantas [69c].
Actividad elicitora del quitosano y sus oligómeros. Aunque el quitosano y la quitina tienen estructuras químicas similares sus actividades elicitoras son diferentes [70]. Muchísimos estudios han demostrado que el quitosano es un poderoso estimulante del sistema de resistencia adquirida a patógenos en plantas. Entre los estudios relacionados con el estímulo de mecanismos de defensa por quitosano se pueden mencionar los siguientes [7]:
Producción de proteínas relacionadas a la patogénesis (origen y evolución de las enfermedades): el quitosano ha sido descrito como un potente elicitor en plantas, estimulando respuestas defensivas como por ejemplo la producción de un amplio espectro de compuestos, proteínas y péptidos con actividad antimicrobiana que las protegen del ataque de patógenos. Las proteínas inducidas por quitosano más conocidas y estudiadas son enzimas hidrolíticas como las quitinasas y β–1,3–glucosanasas, las cuales usualmente actúan sinérgicamente destruyendo las paredes celulares de hongos, que contienen quitina, evitando su crecimiento en las plantas. Entre los cultivos donde se ha comprobado la elicitación por quitosano se encuentran tomate [71], durazno [72], fresa [73], uva de mesa [74], etc. En los dos últimos casos, resulta interesante la proposición de que la estructura no porosa en la cutícula de las frutas podría impedir físicamente el ingreso de quitosano por el tejido, evitando con ello su efecto elicitor en frutas sin heridas. Se presume que el quitosano puede actuar de diferentes formas para aumentar las funciones de los llamados genes relacionados a la patogénesis, incluyendo la activación de receptores en la superficie o en la membrana celular, así como la afectación de los arreglos conformacionales del ADN influenciando así la transcripción genética [75], entre otras. El peso molecular del quitosano parece tener una influencia marcada en este tipo de aplicaciones, como ha sido observado en semilleros de arroz infectados con Magnaporthe grisea, en los cuales hubo una mayor reducción de la infección en las hojas tratadas con los oligómeros de peso molecular 1.335 Da, comparado con el control y con quitosanos de 3.350 y 50.000 Da [76].
Producción de enzimas de defensa: entre las enzimas de defensa que pueden ser inducidas por el quitosano se pueden mencionar la fenilalanina amonio liasa (FAL), la peroxidasa, la polifenol oxidasa (PFO), la catalasa y la superóxido dismutasa (SOD). En el caso de la FAL se ha reportado que la piel de uvas de mesa rociadas con quitosano al 1% presentaron el doble de actividad que las no tratadas y que los tratamiento pre– y post–cosecha redujeron la infección por el moho gris [77]. Ente otros productos donde también se ha observado la inducción de FAL por quitosano se pueden mencionar arroz [78], soya [79], pera [80], mango [81], etc.
Por otro lado, también se ha reportado que luego de aplicar tratamientos con quitosano se ha observado un incremento apreciable en la actividad de (a) la peroxidasa alrededor de las heridas en peras [80]; (b) la PFO en plántulas de arroz [78]; (c) la catalasa en plántulas de maíz [82]. Algunos resultados interesantes han sido reportados recientemente sobre la capacidad del quitosano, en solución y en solución de nanoparticulas, para inducir la expresión de genes relacionados a SOD en hojas de tomate provenientes de plantas infectadas con Fusarium andiyazi, con valores llamativamente mayores para el sistema de nanopartículas (1,2 y 4,54 veces, respectivamente)[83].
Producción de metabolitos secundarios de defensa: se puede considerar que el quitosano logra ser percibido por las plantas como un elicitor biótico de origen fúngico [84]. Existe un sinnúmero de especies vegetales en las cuales se ha probado que el quitosano estimula notoriamente la producción de metabolitos secundarios, los cuales normalmente contribuyen con su sistema de protección. Sin embargo, también se ha planteado que, bajo ciertas condiciones bien determinadas, pudiera aprovecharse este hecho para incrementar la producción de sustancias específicas, como por ejemplo limoneno (un aumento de 17 veces) [85] y menta [86].
En general, se ha encontrado que el tratamiento con quitosano puede inducir distintos mecanismos de defensa en plantas, como por ejemplo la suberización durante el procedo de cicatrización de heridas en tubérculos de papas [87], y también vía estímulo de la producción de metabolitos secundarios, incluyendo: (a) fitoalexinas, como se observó en la acumulación de naftoquinonas antifúngicas en las “jarras” de la planta carnívora Nepenthes khasiana [88]; (b) lignina, como se ha determinado durante el tratamiento del fruto del dragón con dispersiones sub–micrónicas de partículas de quitosano para controlar la antracnosa [89]; (c) compuestos fenólicos, como se ha observado en el caso de las hojas de té luego de tratamientos pre– y post–cosecha con quitosanos de peso molecular 8.000 y 10.000 Da [90]; (d) callosa, cuya deposición juega un rol importante en la limitación del esparcimiento de virus [91] pero parece estar controlada por el peso molecular y el grado de acetilación del quitosano empleado durante el tratamiento [92].
Activación de receptores específicos: aunque son pocos los reportes para receptores específicos de oligómeros de quitosano, se han aislado algunas lectinas con capacidad para unir este tipo de fragmentos a partir de plantas como la zarzamora (Rubus fruticosus) [93] y la col china de hojas sueltas [94]. En el caso de la zarzamora se ha propuesto que sus funciones pudieran ser la inhibición de patógenos que contienen quitosano en sus paredes celulares o iniciar mecanismos de defensa específicos para este tipo de atacantes. Por ello, es necesario realizar más investigaciones sobre este particular que ayuden a clarificar las características, funciones y mecanismos de acción de estos receptores.
Algunas tendencias para el futuro próximo. Como puede apreciarse de esta muy sucinta exposición sobre los efectos beneficiosos y los modos de acción de estos dos biopolímeros en aplicaciones agrícolas, la quitina y el quitosano son materiales que no han perdido actualidad en este campo, luego de casi 60 años de haber comenzado sus aplicaciones en el área de forma sistemática. Por el contrario, éstas se han hecho cada vez más actuales debido a que el desarrollo de sus múltiples formas de aplicación parece correr paralelo al desarrollo de las nano y biotecnologías, como por ejemplo el desarrollo de nanosensores, nanogeles, películas basadas en materiales híbridos, etc., potencialmente útiles en agricultura. Por ello, en esta parte final se presenta una lista de desafíos que han sido emprendidos o pueden emprenderse a corto plazo en el área, especialmente con miras a lograr materiales y tecnologías cada vez más ambientalmente amigables:
Encontrar nuevas fuentes y/o procesos que permitan obtener quitinas/quitosanos más “verdes” aún, como por ejemplo la obtención de oligoquitinas a partir de algas o levaduras [95]. Un beneficio adicional de este desarrollo podría ser la obtención de materiales con propiedades más reproducibles, menos contaminados e independientes de las limitaciones estacionales, que afectan hoy sus principales métodos de producción.
Encontrar nuevos procesos y/o enzimas que generen materiales útiles a partir de residuos quitinosos, como por ejemplo emplear enzimas obtenidas de microorganismos extremófilos como el Thermococcus kodakaraensis KOD1, aislado de una fumarola sulfurosa en Japón y que produce una desaminasa [96]. Este tipo de enzimas pudiera catalizar la reacción: Quitosano à Glucosa + NH3.
Desarrollar nuevos métodos para el seguimiento cinético cuantitativo de la degradación de los materiales quitinosos en suelos. Entre otras cosas, el desarrollo de este tipo de análisis permitiría conocer mejor las implicaciones de la amonificación de la quitina y el quitosano, así como sus derivados, en suelos [97].
Desarrollar estudios sistemáticos de combinaciones de materiales quitinosos con microorganismos productores de volátiles fungicidas, como por ejemplo la investigación adelantada para la combinación quitina/Paenibacillus polymyxa, en la cual se encontró que 7 (benzotiazol, benzaldehído, undecanal, dodecanal, hexadecanal, 2–tridecanona y fenol) de los 42 volátiles generados resultan inhibidores del crecimiento de Fusarium oxysporum [98].
Realizar estudios sistemáticos con quitina y/o quitosano para tratar de favorecer la proliferación de antagonistas que controlen efectivamente fitopatógenos específicos, por ejemplo, estudiar el efecto de quitina y/o quitosano sobre Pseudomonas jessenii, un antagonista reportado para el control de Rizoctonia solani en lechuga [99].
Adelantar trabajos sobre posibles efectos de sustancias similares a los llamados “factores de nodulación”, basados en derivados oligoméricos de quitina y/o quitosano y ácidos grasos [100], incluyendo derivados preparados por interacción iónica del grupo amina protonado con grupos carboxilatos del ácido. Continuar la realización de estudios sistemáticos de nanosistemas para la dosificación de oligosacáridos de quitina y quitosano. El panorama se vislumbra tan amplio que pudiera también incluir estudios de liberación controlada de oligosacáridos de quitosano con nanopartículas de quitosano.
BIBLIOGRAFÍA
[1] Emmott S. “Diez mil millones”. Editorial Anagrama, Colección Argumentos, Barcelona España, 2013
[2] Riechmann J “La revolución (ecosocialista y ecofeminista) tendríamos que haberla hecho ayer”. Revista Theomai, 32, 13 (2015)
[3] Lárez Velásquez C, Millán Barrios E “Chitosan for Pesticide Control on Environmental Protection and Water Purification”. En: “Water Purification”. Chapter 4. Nova Publisher, Series: Air, Water and Soil Pollution Science and Technology (2009). ISBN: 978–1–60741–599–2
[4] Sabadini R, Martins V, Pawlicka A “Synthesis and characterization of gellan gum:chitosan biohydrogels for soil humidity control and fertilizer release”, Cellulose, 22(3), 2045 (2015)
[5] Pereira A, Martins A, Paulino A, Fajardo A, Guilherme M, Iecher Faria M, Linde G, Rubira A, Muniz E “Recent Advances in Designing Hydrogels from Chitin and Chitin–Derivatives and their Impact on Environment and Agriculture: A review”, Rev. Virtual Quim., 9, 17 (2017)
[6] Malerba M, Cerana R “Recent advances of chitosan applications in plants”. Polymers, 10(2), 118 (2018)
[7] Xing K, Zhu X, Peng X, Qin S “Chitosan antimicrobial and eliciting properties for pest control in agriculture: a review”. Agron. Sustain. Dev., 35(2), 569 (2015)
[8] Kong M, Chen X, Xing K, Park H. “Antimicrobial properties of chitosan and mode of action: a state of the art review”. Int. J. Food Microbiol., 144(1), 51 (2010)
[9] Fayaz A, Ao Z, Girilal M, Chen L, Xiao S, Kalaichelvan P, Yao X. “Inactivation of microbial infectiousness by silver nanoparticles–coated condom: a new approach to inhibit HIV–and HSV–transmitted infection”, Int. J. Nanomedicine, 7, 5007 (2012).
[10] Smith D, Yazici S “Chitosan–based nitric oxide donor compositions”. Patent No. US 6,451,337 B1 (2002)
[11] Agrawal S, Pruthi J. “Development and evaluation of matrix type transdermal patch of ethinyl–estradiol and medroxyprogesterone acetate for anti–implantation activity in female Wistar rats”. Contraception, 84(5), 533 (2011)
[12] Smith R “Chitosan as a contraceptive”. Patent No.: US 4,474,769 A (1984)
[13] (a) KTH Royal Institute of Technology. “A next–generation non–hormonal contraceptive for women is being developed in Sweden”. 29 March (2018). Available: https://medicalxpress.com/news/2018–03–next–generation–non–hormonal–contraceptive–women.html; Ultimo acceso 7 de febrero 2019 (b) Kootala S, Filho L, Srivastava V, Linderberg V, Moussa A, Trombotto H, Crouzier T. “Reinforcing mucus barrier properties with low molar mass chitosans”. Biomacromolecules, 19(3), 872 (2018)
[14] Dai T, Tanaka M, Huang Y, Hamblin M. “Chitosan preparations for wounds and burns: Antimicrobial and wound–healing effects”. Exp. Rev. Anti Infect. Ther., 9, 857 (2011)
[15] Izumi R, Komada S, Ochi K, Karasawa L, Osaki T, Murahata Y, Tsuka T, Imagawa T, Itoh N, Okamoto Y, Izawa H, Morimoto M, Saimoto H, Azuma K, Ifuku S. “Favorable effects of superficially deacetylated chitin nanofibrils on the wound healing process”. Carbohydr. Polym., 123, 461 (2015)
[16] Mitchell R, Alexander M “Chitin and the biological control of Fusarium diseases”, Plant Dis. Rep., 45, 487 (1961)
[17] Mitchell R, Alexander M “Microbiological processes associated with the use of chitin for biological control”. Soil Sci. Soc. Am. J., 26, 556 (1962)
[18] Rodriguez–Kabana R. “Organic and inorganic nitogen amendments to soil as nematode suppressants”. J. Nematol., 18, 129 (1986)
[19] Culbreath A, Rodríguez–Kábana R, Morgan–Jones G “The use of hemicellulosic waste matter for reduction of the phytotoxic effects of chitin and control of root–knot nematodes”. Nematropica, 15, 49 (1985)
[20] Tian H, Riggs R, Crippen D. “Control of soybean cyst nematode by chitinolytic bacteria with chitin substrate”. J. Nematology, 32(4), 370 (2000)
[21] Westerdahl B, Carlson H, Grant J, Radewald J, Welch N, Anderson C, Darso J, Kirby D, Shibuya F “Management of plant–parasitic nematodes with a chitin–urea soil amendment and other materials”. J. Nematol., 24, 669 (1992)
[22] Maurer C, Baker R “Chitin–lignin amendments for control of bean root rot”, Phytopathology (Abstr.), 53, 882 (1963)
[23] Culbreath A, Rodriguez–Kabana R, Morgan–Jones G “Chitin and Paecilomyces lilacinus for control of Meloidogyne arenaria”. Nematrópica, 16, 153 (1986)
[24] Lárez–Velásquez C, Rojas Pirela M “Biochemical Aspects of the Chitin Fungicidal Activity in Agricultural Uses” en Chitosan in the Preservation of Agricultural Commodities, Oxford: Academic Press, 2016
[25] Beier S, Bertilsson S “Bacterial chitin degradation—mechanisms and ecophysiological strategies”. Frontiers in Microbiol., 4, Article 149, 12 (2013)
[26] Kumeta Y, Inami Y, Ishimaru K, Yamazaki Y, Sameshima–Saito R, Saito A. “Thermogravimetric evaluation of chitin degradation in soil: implication for the enhancement of ammonification of native organic nitrogen by chitin addition”, Soil Science and Plant Nutrition, 64(4), 512 (2018)
[27] Bell A, Hubbard J, Liu L, Davis R, Subbarao R “Effects of chitin and chitosan on the incidence and severity of Fusarium yellows of celery”. Plant Disease, 82(3), 322 (1998)
[28] Hora T, Baker R “Soil fungistasis: microflora producing a volatile inhibitor”. Trans. British Mycol. Soc., 59(3), 491 (1972)
[29] Schippers B, Palm L. “Ammonia, a fungistatic volatile in chitin–amended soil”. Nether. J. Plant Pathol., 79, 279 (1973)
[30] Gould W, Bryant R, Trofymow J, Anderson R, Elliott E, Coleman D. “Chitin decomposition in a model soil system”. Soil Biol. Biochem., 13, 487 (1981)
[31] Raza W, Yuan J, Ling N, Huang Q, Shen Q “Production of volatile organic compounds by an antagonistic strain Paenibacillus polymyxa WR–2 in the presence of root exudates and organic fertilizer and their antifungal activity against Fusarium oxysporum f. sp. Niveum”. Biological Control, 80, 89 (2015)
[32] Sultana V, Ara J, Parveen G, Ehteshamul–Haque S, Ahmad V. “Role of crustacean chitin, fungicides and fungal antagonist on the effcacy of Pseudomonas aeruginosa in protecting chilli from root rot”, Pak. J. Bot., 38, 1323 (2006)
[33] Patiño L, Bustamante E, Salazar L, «Efecto de sustratos foliares sobre la sigatoka negra (Mycosphaerella fjiensis Morelet) en banano (Musa paradisiaca L.) y plátano (Musa acuminate Colla)». Agricultura Técnica, 67, 437 (2007)
[34] Roberts W, Selitrennikoff C. “Plant and bacterial chitinases differ in antifungal activity”. J. Gen. Microbiol., 134, 169 (1988)
[35] Vaidya R, Shah I, Vyas P, Chhatpar H “Production of chitinase and its optimization from a novel isolate Alcaligenes xylosoxydans: potential in antifungal biocontrol”. World J. Microb. Biotechnol., 17, 691 (2001)
[36] Singh N, Verma R, Shanmugam V. “Extracellular chitinases of Fuorescent pseudomonads antifungal to Fusarium oxysporum f. sp. dianthi causing carnation wilt”. Curr. Microbiol., 52, 310 (2006)
[37] Malathi P, Viswanathan R. “Role of microbial chitinase in the biocontrol of sugarcane red rot caused by Colletotrichum falcatum Went”. Eur. J. Biol. Sci., 6, 17 (2013)
[38] San–Lang W, Yieh T, Shih I. “Purifcation and characterization of a new antifungal compound produced by Pseudomonas aeruginosa K–187 in a shrimp and crab shell powder medium”. Enz. Microb. Technol., 25, 439 (1999)
[39] San–Lang W, Ing–Lung S, Chuan–Lu W, Tseng K, Wen–Teish C, Twu Y, Ro J, Wang C. “Production of antifungal compounds from chitin by Bacillus subtilis”. Enz. Microb. Technol., 31, 321 (2002)
[40] Murphy J, Rafferty S, Cassells A. “Stimulation of wild strawberry (Fragaria vesca) arbuscular mycorrhizas by addition of shellfish waste to the growth substrate: interaction between mycorrhization, substrate amendment and susceptibility to red core (Phytophthora fragariae)”. App. Soil Ecol., 15, 153 (2000)
[41] Staehelin C, Schultze M, Tokuyasu K, Poinsot V, Promé J, Kondorosi E, Kondorosi A. “N–deacetylation of Sinorhizobium meliloti Nod factors increases their stability in the Medicago sativa rhizosphere and decreases their biological activity”. Mol. Plant–Microbe Interact., 13, 72 (2000)
[42] Long S. “Rhizobium symbiosis: Nod factors in perspective”. Plant Cell, 8, 1885 (1996)
[43] Lárez–Velásquez C, Rivas A, Velásquez W, Bahsas A “Amidación del quitosano con cloruro de oleoilo”. Rev. Iberoam. Polím., 8(4), 229 (2007)
[44] Gryndler M, Jansa J, Hršelová H, Chvátalová I, Vosátka M. “Chitin stimulates development and sporulation of arbuscular mycorrhizal fungi”. App. Soil Ecol., 22, 283 (2003)
[45] Millet Y, Danna C, Clay N, Songnuan W, Simon M, Werck–Reichhart D, Ausubel F. “Innate immune responses activated in Arabidopsis roots by microbe–associated molecular patterns”, Plant Cell., 22, 973 (2010)
[46] Kurosaki F, Tashiro N, Nishi A. “Role of chitinase and chitin oligosaccharides in lignifcation response of cultured carrot cells treated with mycelial walls”. Plant Cell Physiol., 29, 527 (1988)
[47] Barber M, Bertram R, Ride J. “Chitin oligosaccharides elicit lignification in wounded wheat leaves”. Physiol. Mol. Plant Pathol., 34, 3 (1989)
[48] Jung W, Mabood F, Souleimanov A, Smith D. “Effect of chitin hexamer and Thuricin 17 on lignifcation–relate and antioxidative enzymes in soybean plants”. J. Plant Biol., 51, 145 (2008)
[49] Tanabe S, Okada M, Jikumaru Y, Yamane H, Kaku H, Shibuya N, Minami E. “Induction of resistance against rice blast fungus in rice plants treated with a potent elicitor, N–acetylchitooligosaccharide”. Biosci, Biotechnol, Biochem., 70, 1599 (2006)
[50] Kuchitsu K, Kosaka H, Shiga T, Shibuya N. “EPR evidence for generation of hydroxyl radical triggered by N–acetylchitooligosaccharide elicitor and a protein phosphatase inhibitor in suspension–cultured rice cells”. Protoplasma, 188, 138 (1995)
[51] Wan J, Zhang X, Neece D, Ramonell K, Clough S, Kim S, Stacey M, Stacey G “A LysM receptor–like kinase plays a critical role in chitin signaling and fungal resistance in Arabidopsis”. Plant Cell., 20, 471 (2008)
[52] Peumans W, De Ley M, Broekaert W. “An unusual lectin from stinging nettle (Urtica dioica) rhizomes”. Fed. Eur. Biochem. Soc., 177(1), 99 (1983)
[53] Broekaert W, Van Parijs J, Leyns F, Joos H, Peumans W. “A chitin–binding lectin from stinging nettle rhizomes with antifungal properties”. Science (New Series), 245, 1100 (1989)
[54] Felix G, Regenass M, Boller T. “Specifc perception of subnanomolar concentrations of chitin fragments by tomato cells: induction of extracellular alkalinization, changes in protein phosphorylation”. Plant J., 4(2), 307 (1993)
[55] (a) Shibuya N, Kaku H, Kuchitsu K, Maliarik M. “Identification of a novel high affinity binding site for N–acetylchitooligosaccharide elicitor in the membrane fraction from suspension–cultured rice cells”. Federation of European Biochemical Societies Letters, 329(1–2), 75 (1993); (b) Ito Y, Kaku H, Shibuya H. “Identification of a high–affnity binding protein for N–acetylchitooligosaccharide elicitor in the plasma membrane of suspension–cultured rice cells by affnity labeling”. Plant J., 12, 347 (1997); (c) Kaku H, Nishizawa Y, Ishii–Minami N, Akimoto–Tomiyama C, Dohmae N, Takio K, Minam E, Shibuya N. “Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor”. Proceed. Nat. Acad. Sci., 103(29), 11086 (2006)
[56] Miya A, Albert P, Shinya T, Desaki Y, Ichimura K, Narusaka Y, Kawakami N, Kaku N, Shibuya N. “CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis”. Proc. Nat. Acad. Sci., 104, 19613 (2007)
[57] Sánchez–Vallet A, Saleem–Batcha R, Kombrink A, Hansen G, Valkenburg D, Thomma B, Mesters J. “Fungal effector Ecp6 outcompetes host immune receptor for chitin binding through intrachain LysM dimerization”. eLife, 2, e00790 (2013)
[58] Ohta K, Morishita S, Suda K, Kobayashi N, Hosoki T. “Effects of chitosan soil mixture treatment in the seedling stage on the growth and flowering of several ornamental plants”, J. Japanese Soc. Hort. Sci., 73(1), 66 (2004)
[59] Guan Y, Hu J, Wang X, Shao C. “Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress”. J. Zhejiang University Science B, 10(6), 427 (2009)
[60] Lárez–Velásquez C, Chirinos A, Tacoronte M, Mora A. “Chitosan oligomers as bio–stimulants to zucchini (Cucurbita pepo) seeds germination”. Agriculture (Poľnohospodárstvo), 5(8), 113 (2012)
[61] Chirinos A, Tacoronte M, Rojas A, Lárez–Velásquez C. “Estudios preliminares del efecto del quitosano en la germinación y protección en semillas de calabacín (Cucurbita pepo)«. Póster en VII Escuela Internacional de Polímeros, Margarita, Venezuela (2010)
[62] Wieczorek A, Hetz S, Kolb S. “Microbial responses to chitin and chitosan in oxic and anoxic agricultural soil slurries”. Biogeosciences, 11, 3339–3352 (2014)
[63] Mihara S “Change in glucosamine content of Chlorella cells during their life cycle”. Plant Cell Physiol., 2, 25 (1961)
[64] Bartnicki–Garcia S, Nickerson W. “Isolation, composition and structure of cell walls of filamentous and yeast–like forms of Mucor rouxii”. Biochim. Biophys. Acta, 58, 102 (1962)
[65] Sawaguchi A, Ono S, Oomura M, Inami K, Kumeta Y, Honda K, Sameshima–Saito R, Sakamoto K, Ando A, Saito A. “Chitosan degradation and associated changes in bacterial community structures in two contrasting soils”. Soil Science and Plant Nutrition, 61(3), 471 (2015)
[66] Postma J, Stevens L, Wiegers G, Davelaar E, Nijhuis E. “Biological control of Pythium aphanidermatum in cucumber with a combined application of Lysobacter enzymogenes strain 3.1T8 and chitosan”. Biological Control, 48, 301 (2009)
[67] Kaur K, Dattajirao V, Shrivastava V, Bhardwaj U. “Isolation and characterization of chitosan–producing bacteria from beaches of Chennai, India”. Enzyme research, Article ID 421683, 6 pages (2012)
[68] Hsu S, Chung Y, Chang C, Sung H. “Purification and Characterization of Two Chitosanase Isoforms from the Sheaths of Bamboo Shoots”. J. Agri. Food Chem., 60(2), 649 (2012)
[69] (a) Lerouge P, Roche P, Faucher C, Maillet F, Truchet G, Promé J, Dénarié J. “Symbiotic host–specificity of Rhizobium meliloti is determined by a sulphated and acylated glucosamine oligosaccharide signal”. Nature, 344, 781 (1990); (b) Sahur A. “The Interaction between Endophytic Actinomycetes and Rhizobium in Leguminous Plants”. J. Trop. Crop Sci., 2(3), 29 (2015); (c) Limpens E, van Zeijl A, Geurts R. “Lipochitooligosaccharides modulate plant host immunity to enable endosymbioses”. Ann. Rev. Phytopathology, 53, 311 (2015)
[70] Yin H, Du Y, Dong Z. “Chitin Oligosaccharide and Chitosan Oligosaccharide: Two Similar but Different Plant Elicitors”. Front. Plant Sci., 7, Article 522, 4 pages (2016)
[71] Benhamou N, Lafontaine P, Nicole M. “Induction of systemic resistance to Fusarium crown and root rot in tomato plants by seed treatment with chitosan”. Phytopathology, 84(12), 1432 (1994)
[72] Ma Z, Yang L, Yan H, Kennedy J, Meng X. “Chitosan and oligochitosan enhance the resistance of peach fruit to brown rot”. Carbohydr. Polym., 94, 272 (2013)
[73] El–Ghaouth A, Arul J, Grenier J, Asselin A. “Antifungal activity of chitosan on two post–harvest pathogens of strawberry fruits”. Phytopathology, 82, 398 (1992)
[74] Romanazzi G, Gabler F, Margosan D, Mackey D, Smilanick J. “Effect of chitosan dissolved in different acids on its ability to control postharvest gray mold of table grape”. Phytopathology, 99, 1028 (2009)
[75] Hadwiger L “Host–parasite interactions: elicitation of defense responses in plants with chitosan”. EXS, 87, 185 (1999)
[76] Lin W, Hu X, Zhang W, Rogers W, Cai W. “Hydrogen peroxide mediates defence responses induced by chitosans of different molecular weights in rice”. J. Plant Physiol., 162, 937 (2005)
[77] Romanazzi G, Nigro F, Ippolito A, Di Venere D, Salerno M “Effects of pre– and postharvest chitosan treatments to control storage grey mold of table grapes”. J. Food Sci., 67, 1862 (2002)
[78] Li B, Liu B, Shan C, Ibrahim M, Lou Y, Wang Y, Xie G, Li H, Sun G “Antibacterial activity of two chitosan solutions and their effect on rice bacterial leaf blight and leaf streak”. Pest Manag. Sci., 69, 312 (2013)
[79] Chen H, Seguin P, Archambault A, Constan L, Jabaji S. “Gene expression and isoflavone concentrations in soybean sprouts treated with chitosan”. Crop Sci., 49, 224 (2009)
[80] Meng X, Yang L, Kennedy J, Tian S. “Effects of chitosan and oligochitosan on growth of two fungal pathogens and physiological properties in pear fruit”. Carbohydr. Polym., 81, 70 (2010)
[81] Berumen–Varela G, Coronado–Partida D, Ochoa–Jiménez V, Chacón–López A, Gutiérrez–Martínez P. “Effect of chitosan on the induction of disease resistance against Colletotrichum sp. in mango (Mangifera indica L) cv”. Tommy Atkins, 66, 16 (2015)
[82] Guan Y, Hu J, Wang X, Shao C. “Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress”. J. Zhejiang Univ. Sci., B10, 427 (2009)
[83] Chun S, Chandrasekaran M. “Chitosan and chitosan nanoparticles induced expression of pathogenesisrelated proteins genes enhances biotic stress tolerance in tomato”. Int. J. Biol. Macromol., 125, 948 (2019)
[84] Narayani M, Srivastava S. “Elicitation: a stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production”. Phytochem. Rev., 16, 1227 (2017)
[85] Lockwood G, Bunrathep S, Songsak T, Ruangrungsi N. “Production of d–Limonene in Chitosan Elicited Citrus japonica Suspension Cultures”. J. Essential Oils Res., XX, 113 (2007)
[86] Chang J, Shin J, Chung I, Lee H. “Improved menthol production from chitosan–elicited suspension culture of Mentha piperita”. Biotechnology Letters, 20, 1097 (1998)
[87] Ozeretskovskaia O, Vasiukova N, Chalenko G, Gerasimova N, Revina T, Valueva T. “Wound healing and induced resistance in potato tubers”. Prikl. Biokhim. Mikrobiol., 45, 220 (2009)
[88] Eilenberg H, Pnini–Cohen S, Rahamim Y, Sionov E, Segal E, Carmeli S, Zilberstein A. “Induced production of antifungal naphthoquinones in the pitchers of the carnivorous plant Nepenthes khasiana”. J. Exp. Bot., 61, 911 (2010)
[89] Ali A, Zahid N, Manickam S, Siddiqui Y, Alderson P, Maqbool M “Induction of lignin and pathogenesis related proteins in dragon fruit plants in response to submicron chitosan dispersions”, Crop Protection, 63, 83 (2014)
[90] Srisornkompon P, Pichyangkura R, Chadchawan S. “Chitosan Increased Phenolic Compound Contents in Tea (Camellia sinensis) Leaves by Pre– and Post–Treatments”, J. Chitin and Chitosan Science, 2, 1 (2014)
[91] Levy A, Guenoune–Gelbart D, Epel D. “β–13–Glucanases Plasmodesmal gate keepers for intercellular communication”. Plant Signal Behav., 2, 288 (2007)
[92] Kauss H, Jeblick W, Domard A. “The degree of polymerization and N–acetylation of chitosan determine its ability to elicit callose formation in suspension–cultured cells and protoplasts of Catharanthus roseus”. Planta, 178, 385 (1989)
[93] Liénart Y, Gautier C, Domard A “Isolation from Rubus cell–suspension cultures of a lectin specific for glucosamine oligomers”. Planta, 184(1), 8 (1991)
[94] Chen H, Xu L. “Isolation and Characterization of a Novel Chitosan–Binding Protein from Non–Heading Chinese Cabbage Leaves”. J. Integrative Plant Biology, 47(4), 452 (2005)
[95] De Ros D, Cerro M, Canovas E, Turk O, Yebra J, Roca J, et al. “Chitin and chitosan producing methods”. U.S. Patent Application No. 15/537,247. 30 Nov. 2017
[96] Anaka T, Takahashi T, Fukui T, Fujiwara S, Atomi H, Imanaka T. “Characterization of a novel glucosamine–6–phosphate deaminase from a hyperthermophilic archaeon”. J. Bacteriol., 187(20), 7038 (2005)
[97] Kumeta Y, Inami Y, Ishimaru K, Yamazaki Y, Sameshima–Saito R, Saito A “Thermogravimetric evaluation of chitin degradation in soil: implication for the enhancement of ammonification of native organic nitrogen by chitin addition”. Soil Science and Plant Nutrition, 64(4), 512 (2018)
[98] Raza W, Yuan J, Ling N, Huang Q, Shen Q. “Production of volatile organic compounds by an antagonistic strain Paenibacillus polymyxa WR–2 in the presence of root exudates and organic fertilizer and their antifungal activity against Fusarium oxysporum f. sp. Niveum”. Biological Control, 80, 89 (2015)
[99] Adesina M, Grosch R, Lembke A, Vatchev T, Smalla K “In vitro antagonists of Rhizoctonia solani tested on lettuce: rhizosphere competence, biocontrol e⁄ciencyand rhizosphere microbial community response”, FEMS Microbiol. Ecol., 69, 62 (2009)
[100] Berthelot N, Brossay A, Gasciolli V, Bono J, Baron A, Beau J, Urban D, Boyer F, Vauzeilles B. “Synthesis of lipo–chitooligosaccharide analogues and their interaction with LYR3, a high affinity binding protein for Nod factors and Myc–LCOs”, Org. Biomol. Chem., 15(37), 7802 (2017)
Imagen de portada:
«Bread»by Jerzy Durczak is licensed under CC BY-NC 2.0![]()
![]()
![]()

Excelente artículo!!
Me gustaMe gusta